


説明
Hieff Trans™リポソームトランスフェクション試薬は、DNA、RNA、オリゴヌクレオチドのトランスフェクションに適した汎用性の高いリポソームトランスフェクション試薬で、ほとんどの真核細胞に対して高いトランスフェクション効率を実現します。独自の配合により培地に直接添加でき、血清の存在がトランスフェクション効率に影響を与えないため、血清除去による細胞へのダメージを軽減します。トランスフェクション後、核酸-Hieff Trans™複合体を除去したり、新しい培地と交換したりする必要がなく、4~6時間後に除去することも可能です。
Hieff Trans™は滅菌液状で提供されます。通常、24ウェルプレートの場合、1回あたり約1.5μLのHieff Trans™ 1mLで約660回のトランスフェクションが可能です。6ウェルプレートの場合、1回あたり約6μLのHieff Trans™ 1mLで約660回のトランスフェクションが可能です。160回のトランスフェクションが可能です。
特徴
- 優れた効率:一過性トランスフェクションや安定的トランスフェクションを含む、幅広い細胞株で優れたトランスフェクション性能を発揮します。
- 幅広い適応性:さまざまな細胞株における優れたトランスフェクション効率と高レベルの組み換えタンパク質発現
- 低毒性:YEASENのトランスフェクション試薬は、多様な細胞の活動にほとんど影響を与えません。
- 簡単な操作:血清存在下での効果が実証されており、トランスフェクション後の培地交換が不要です。
- コスト効率:より手頃な価格で競争力のあるトランスフェクション効果
応用
- 細胞トランスフェクション
仕様
| 形状 | 液体 |
| 血清適合性 | はい |
| 細胞の種類 | 樹立細胞株 |
| サンプルタイプ | プラスミドDNA、合成siRNA |
| トランスフェクション技術 | 脂質ベースのトランスフェクション |
コンポーネント
| コンポーネント番号 | 名前 | 40802ES02 | 40802ES03 | 40802ES08 |
| 40802 | Hieff Trans ™リポソームトランスフェクション試薬 | 0.5 mL | 1mL | 5×1mL |
配送と保管
本製品は保冷剤入りで、2~8℃で1年間保存できます。冷凍しないでください。
数字

引用元: Communications Biology 第5巻、記事番号:1296(2022年)
- トランスフェクション効果の実証
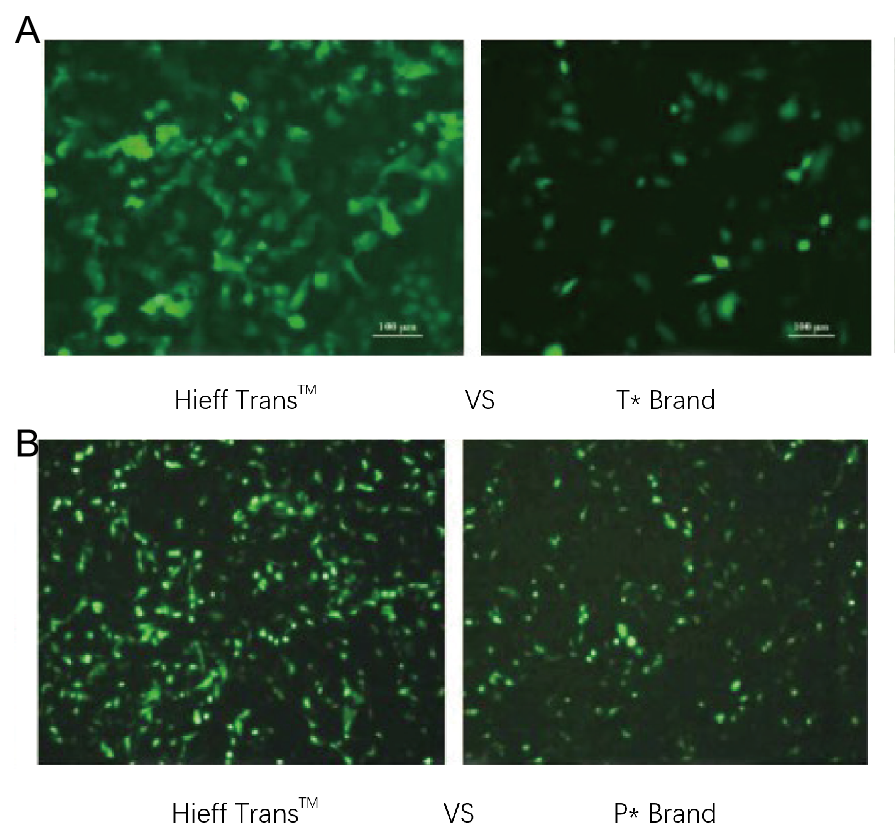
図 1. Hieff Trans™ リポソームトランスフェクション試薬は、競合ブランドのトランスフェクション試薬よりも優れています(図 1a の T* ブランド、図 1b の P* ブランド)。
各試薬を用いて、96ウェルフォーマットで標的細胞株(図1aはHela細胞株、図1bはDF-1細胞株)にトランスフェクションを行いました。トランスフェクション後48時間でGFP発現を解析しました。Hieff Trans™ Liposome Transfection Reagentは、競合製品と比較して高いGFPトランスフェクション効率を示しました。
- 検証済み細胞株
| 製品名 | Hieff TransTMリポソームトランスフェクション試薬 | |||||
| 検証済み細胞株 | 293T | カル1 | HEK293 | HO1980 | N2A | TS |
| 293F | CHO-K1 | HEK293T | HUVEC | NCI-H1975 | U-87 | |
| 293フィート | COS-7 | ヘラ | MCF10A | NIH-3T3 | ベロ | |
| 3T3 | DF-1 | 肝細胞2C | MCF-7 | ニューロ2a | ウェヒ | |
| A549 | H1299 | 肝細胞3B | MDA-MB-231 | PC-12 | WRL-68 | |
| BV-2 | H520 | ヘパ1-6 | MDA-MB-231-LM2-4175 | 生264.7 | ||
| B50 | ハキャット | 肝G2 | MDCK | SGC-7901 | ||
| C2C12 | HCT116 | HK2 | 外務省 | T47D | … | |
図 2. Hieff Trans™ リポソームトランスフェクション試薬で正常にトランスフェクトされた細胞株のリスト (継続的に更新中)。

「人工光スイッチングRNA結合タンパク質を用いたRNA機能および代謝の光遺伝学的制御。Nat Biotechnol. 2022年1月3日 doi: 10.1038/s41587-021-01112-1」より引用

「損傷したクロマチンへのBRCA1-BARD1複合体のリクルートメントに関する構造的知見。Mol Cell. 2021年7月1日;81(13):2765-2777.e6. doi: 10.1016/j.molcel.2021.05.010」より引用。
「UBQLN2-HSP70軸はポリグリシン-アラニン凝集体を減少させ、C9ORF72動物モデルの行動障害を軽減する。Neuron. 2021年6月16日;109(12):1949-1962.e6. doi: 10.1016/j.neuron.2021.04.023」より引用。
よくある質問:
Hieff Trans™ リポソームトランスフェクション試薬に関するよくある質問
(1)Q:核酸トランスフェクション試薬複合体を調製する際に血清を存在させてもよいですか?
A: 血清の存在はリポソームの形成に影響を与えます。核酸トランスフェクション試薬複合体を調製する際は、無血清培地(通常はMEM培地)の使用をお勧めします。
(2)Q:トランスフェクション試薬は凍結できますか?
A: いいえ。この試薬は 2 ~ 8 ℃ で保管する必要があり、キャップを長期間繰り返し開けないように注意してください。キャップを長期間開けるとリポソームが酸化され、トランスフェクション効率に影響を及ぼします。
(3)Q:Hieff Trans™リポソーム核酸トランスフェクション試薬を使用する際に注意すべきことは何ですか?
A: 1) トランスフェクション操作中、細胞合流度が 80%~95% に達することが望ましく、具体的な播種密度は細胞の状況に応じて決定されます。
2) 高純度 DNA を使用すると、より高いトランスフェクション効率が得られます。
3) トランスフェクション複合体を調製する際には、DNA およびトランスフェクション試薬を無血清培地で希釈する必要があります。
4) トランスフェクション中に培地に抗生物質を添加することはできません。
5) 初回使用時には、最大のトランスフェクション効率を得るために、DNA濃度とカチオン性リポソーム試薬の量を最適化する必要があります。DNAとトランスフェクション試薬の比率は、 一般的には1:2~1:3が推奨されます。
(4)Q:トランスフェクション後に停止させる必要がありますか?
A: 必要ありません。リポソーム複合体は6時間安定です。トランスフェクション前に細胞培地を交換していない場合は、正常な細胞増殖に必要な栄養素を確保するために、4~6時間後に新しい培地に交換する必要があります。ただし、トランスフェクション前に培地を交換している場合は、リポソームトランスフェクション後に培地を交換する必要はありません。
(5)Q:トランスフェクション効率を向上させたい場合、どのような点に注意すればよいですか?
A: a: トランスフェクション時の細胞密度は 90%~95% です。
b: トランスフェクション中は、核酸およびリポソームの希釈に MEM 無血清培地を使用します。
c: トランスフェクション後4〜6時間で培地を交換できます。
(6)Q:DNAとsiRNAの同時トランスフェクションは可能ですか?効果はどうですか?
A: 同時トランスフェクションは可能ですが、別々にトランスフェクションすることをお勧めします。DNAトランスフェクションはsiRNAの6時間後に行う必要があります。同時操作を行うと、siRNAトランスフェクション効率が低下します。
(7)Q:トランスフェクション試薬はレンチウイルスパッケージングトランスフェクションに使用できますか?
A: レンチウイルスのパッケージングは可能ですが、レンチウイルスのパッケージングの効率は必ずしもトランスフェクションの効率に関係しているわけではなく、パッケージングプラスミドの選択とプラスミド間の比率にも関係しています。
(8)Q:Hieff Trans™リポソーム核酸トランスフェクション試薬は懸濁細胞のトランスフェクションに使用できますか?
A: Hieff Trans™ リポソーム核酸トランスフェクション試薬は浮遊細胞へのトランスフェクションに使用できます。詳細はプロトコルをご覧ください。また、浮遊細胞専用のトランスフェクション試薬(カタログ番号 40805、浮遊細胞用リポソーム核酸トランスフェクション試薬)も発売しております。
引用と参考文献:
[1] Liu R, Yang J, Yao J, Zhao Z, He W, Su N, Zhang Z, Zhang C, Zhang Z, Cai H, Zhu L, Zhao Y, Quan S, Chen X, Yang Y. 人工光スイッチングRNA結合タンパク質を用いたRNA機能および代謝の光遺伝学的制御. Nat Biotechnol. 2022年1月3日. doi: 10.1038/s41587-021-01112-1. 電子出版、印刷前. PMID: 34980910. (IF:54.908)
[2] Zhou J, Chen P, Wang H, Liu H, Li Y, Zhang Y, Wu Y, Paek C, Sun Z, Lei J, Yin L. 厳密なPAM認識によりゲノム全体のオフターゲット効果を低減するように設計されたCas12aバリアント. Mol Ther. 2022年1月5日;30(1):244-255. doi: 10.1016/j.ymthe.2021.10.010. Epub 2021年10月20日. PMID: 34687846; PMCID: PMC8753454. (IF:11.454)
[3] Chen S, Cao X, Zhang J, Wu W, Zhang B, Zhao F. circVAMP3はCAPRIN1の相分離を促進し、c-Mycの翻訳を抑制することで肝細胞癌の増殖を抑制する. Adv Sci (Weinh). 2022年3月;9(8):e2103817. doi: 10.1002/advs.202103817. Epub 2022年1月24日. PMID: 35072355; PMCID: PMC8922094. (IF:16.808)
[4] Zhang Y, Yu X, Sun R, Min J, Tang X, Lin Z, Xie S, Li X, Lu S, Tian Z, Gu C, Teng L, Yang Y. スプライシング因子アルギニン/セリンリッチ8は、CACYBPの選択的スプライシングとエクソソームを介した細胞間コミュニケーションを介して、多発性骨髄腫の悪性度と骨病変を促進する. Clin Transl Med. 2022年2月;12(2):e684. doi: 10.1002/ctm2.684. PMID: 35184390. (IF:11.492)
[5] Tang X、Deng Z、Ding P、Qiang W、Lu Y、Gao S、Hu Y、Yang Y、Du J、Gu C。circHNRNPU によってコードされる新規タンパク質は、骨髄微小環境と選択的スプライシングを調節することによって多発性骨髄腫の進行を促進します。 J Exp Clin Cancer Res. 2022 年 3 月 8 日;41(1):85。土井: 10.1186/s13046-022-02276-7。 PMID: 35260179 (IF:11.161)
[6] Hua Z, Wei R, Guo M, Lin Z, Yu X, Li X, Gu C, Yang Y. YTHDF2はSTAT5A/MAP2K2/p-ERK軸を介して多発性骨髄腫細胞の増殖を促進する. Oncogene. 2022年3月;41(10):1482-1491. doi: 10.1038/s41388-022-02191-3. Epub 2022年1月24日. PMID: 35075244. (IF:9.867)
[7] Liang Y, Lu Q, Li W, Zhang D, Zhang F, Zou Q, Chen L, Tong Y, Liu M, Wang S, Li W, Ren X, Xu P, Yang Z, Dong S, Zhang B, Huang Y, Li D, Wang H, Yu W. NamiRNAネットワークを介したエンハンサースイッチングによる乳がんにおける腫瘍抑制因子の再活性化. Nucleic Acids Res. 2021年9月7日;49(15):8556-8572. doi: 10.1093/nar/gkab626. PMID: 34329471; PMCID: PMC8421228. (IF:16.9)
[8] Dai L, Dai Y, Han J, Huang Y, Wang L, Huang J, Zhou Z. 損傷したクロマチンへのBRCA1-BARD1複合体のリクルートメントに関する構造的知見. Mol Cell. 2021年7月1日;81(13):2765-2777.e6. doi: 10.1016/j.molcel.2021.05.010. Epub 2021年6月7日. PMID: 34102105. (IF:17.97)
[9] Zhang K, Wang A, Zhong K, Qi S, Wei C, Shu X, Tu WY, Xu W, Xia C, Xiao Y, Chen A, Bai L, Zhang J, Luo B, Wang W, Shen C. UBQLN2-HSP70軸はポリグリシン-アラニン凝集体を減少させ、C9ORF72動物モデルの行動障害を軽減する. Neuron. 2021年6月16日;109(12):1949-1962.e6. doi: 10.1016/j.neuron.2021.04.023. Epub 2021年5月14日. PMID: 33991504. (IF:17.17)
[10] Liang Y, Lu Q, Li W, Zhang D, Zhang F, Zou Q, Chen L, Tong Y, Liu M, Wang S, Li W, Ren X, Xu P, Yang Z, Dong S, Zhang B, Huang Y, Li D, Wang H, Yu W. NamiRNAネットワークを介したエンハンサースイッチングによる乳がんにおける腫瘍抑制因子の再活性化. Nucleic Acids Res. 2021年9月7日;49(15):8556-8572. doi: 10.1093/nar/gkab626. PMID: 34329471; PMCID: PMC8421228. (IF:16.9)
[11] Li T, Chen X, Qian Y, Shao J, Li X, Liu S, Zhu L, Zhao Y, Ye H, Yang Y. マウスの血糖恒常性維持を可能にするパルス状遺伝子発現のための合成BRETベース光遺伝学デバイス. Nat Commun. 2021年1月27日;12(1):615. doi: 10.1038/s41467-021-20913-1. PMID: 33504786; PMCID: PMC7840992. (IF:14.92)
[12] Pan Y, He X, Li C, Li Y, Li W, Zhang H, Wang Y, Zhou G, Yang J, Li J, Qu J, Wang H, Gao Z, Shen Y, Li T, Hu H, Ma H. 神経活動がCRTC1/CREB軸をリクルートし、転写依存性オートファジーを促進してLTD後期を維持する。Cell Rep. 2021年7月20日;36(3):109398. doi: 10.1016/j.celrep.2021.109398. PMID: 34289350. (IF:9.42)
[13] Liu H, Xing R, Ou Z, Zhao J, Hong G, Zhao TJ, Han Y, Chen Y. Gタンパク質共役受容体GPR17はポリコーム抑制複合体1を介した活性酸素産生を増加させることで神経膠腫の発生を抑制する. Cell Death Dis. 2021年6月12日;12(6):610. doi: 10.1038/s41419-021-03897-0. PMID: 34120140; PMCID: PMC8197764. (IF:8.463)
[14] Fan Y, Wang J, Jin W, Sun Y, Xu Y, Wang Y, Liang X, Su D. CircNR3C2はトリプルネガティブ乳がんにおいてmiR-513a-3pのスポンジングを介してHRD1を介した腫瘍抑制効果を促進する. Mol Cancer. 2021年2月2日;20(1):25. doi: 10.1186/s12943-021-01321-x. PMID: 33530981; PMCID: PMC7851937. (IF:27.403)
[15] Gu C, Wang Y, Zhang L, Qiao L, Sun S, Shao M, Tang X, Ding P, Tang C, Cao Y, Zhou Y, Guo M, Wei R, Li N, Xiao Y, Duan J, Yang Y. AHSA1は多発性骨髄腫における細胞増殖およびプロテアソーム阻害剤耐性に対する有望な治療標的である. J Exp Clin Cancer Res. 2022年1月6日;41(1):11. doi: 10.1186/s13046-021-02220-1. PMID: 34991674; PMCID: PMC8734095. (IF:11.161)
[16] Luo Q, Wu X, Zhao P, Nan Y, Chang W, Zhu X, Su D, Liu Z. OTUD1はAIFの核移行とMCL1の分解を促進することでカスパーゼ非依存性およびカスパーゼ依存性アポトーシスを活性化する. Adv Sci (Weinh). 2021年2月8日;8(8):2002874. doi: 10.1002/advs.202002874. PMID: 33898171; PMCID: PMC8061361. (IF:15.84)
[17] Luo Q, Wu X, Nan Y, Chang W, Zhao P, Zhang Y, Su D, Liu Z. TRIM32/USP11はARID1Aの安定性と扁平上皮癌の発癌性/腫瘍抑制状態をバランスさせる. Cell Rep. 2020年1月7日;30(1):98-111.e5. doi: 10.1016/j.celrep.2019.12.017. PMID: 31914402. (IF:9.42)
[18] Sun X、Peng X、Cao Y、Zhou Y、Sun Y。ADNP は、Wnt/β-カテニンシグナル伝達を調節することによって神経分化を促進します。ナットコミューン。 2020 6 12;11(1):2984。土井: 10.1038/s41467-020-16799-0。 PMID: 32533114; PMCID: PMC7293280。 (IF:14.911)
[19] Yang X, Wang H, Xie E, Tang B, Mu Q, Song Z, Chen J, Wang F, Min J. ERBB3およびERKシグナル伝達の再配線は、ERBB3-E928G変異を有する消化管癌においてFGFR1阻害に対する抵抗性を付与する。Protein Cell. 2020年12月;11(12):915-920. doi: 10.1007/s13238-020-00749-z. PMID: 32632529; PMCID: PMC7719122. (IF:14.872)
[20] Chen, T., Chen, Y., Chen, H. et al. 低発現マイクロRNAの細胞内イメージングのためのデュアル酵素推進非結合DNAウォーキングナノマシン. Nano Res. 12, 1055–1060 (2019). https://doi.org/10.1007/s12274-019-2344-5 (IF:8.21)
[21] Zou Y, Wang A, Shi M, Chen X, Liu R, Li T, Zhang C, Zhang Z, Zhu L, Ju Z, Loscalzo J, Yang Y, Zhao Y. 遺伝子組み換え蛍光センサーを用いた生細胞および生体内における酸化還元ランドスケープとダイナミクスの解析. Nat Protoc. 2018年10月;13(10):2362-2386. doi: 10.1038/s41596-018-0042-5. PMID: 30258175; PMCID: PMC6714056. (IF:13.49)
[22] Zhang K, Zhao X, Chen X, Wei Y, Du W, Wang Y, Liu L, Zhao W, Han Z, Kong D, Zhao Q, Guo Z, Han Z, Liu N, Ma F, Li Z. 後肢虚血治療における注入型ハイドロゲルを用いた間葉系幹細胞由来エクソソームの治療効果の増強. ACS Appl Mater Interfaces. 2018年9月12日;10(36):30081-30091. doi: 10.1021/acsami.8b08449. Epub 2018年8月29日. PMID: 30118197. (IF:8.09)
[23] Hao H, Hu S, Chen H, Bu D, Zhu L, Xu C, Chu F, Huo X, Tang Y, Sun X, Ding BS, Liu DP, Hu S, Wang M. 心筋梗塞後の血管恒常性と心臓リモデリングを阻害する内皮CXCR7の喪失:心血管疾患創薬への示唆. Circulation. 2017年3月28日;135(13):1253-1264. doi: 10.1161/CIRCULATIONAHA.116.023027. Epub 2017年2月2日. PMID: 28154007. (IF:18.881)
文書:
支払いとセキュリティ
お支払い情報は安全に処理されます。 クレジットカードの詳細を保存したり、クレジットカード情報にアクセスすることはありません
あなたも好きかもしれません
問い合わせ
よくある質問
この製品は研究目的のみに使用され、人間や動物の治療や診断に使用することを意図したものではありません。製品とコンテンツは、
特定のアプリケーションでは、追加のサードパーティの知的財産権が必要になる場合があります。